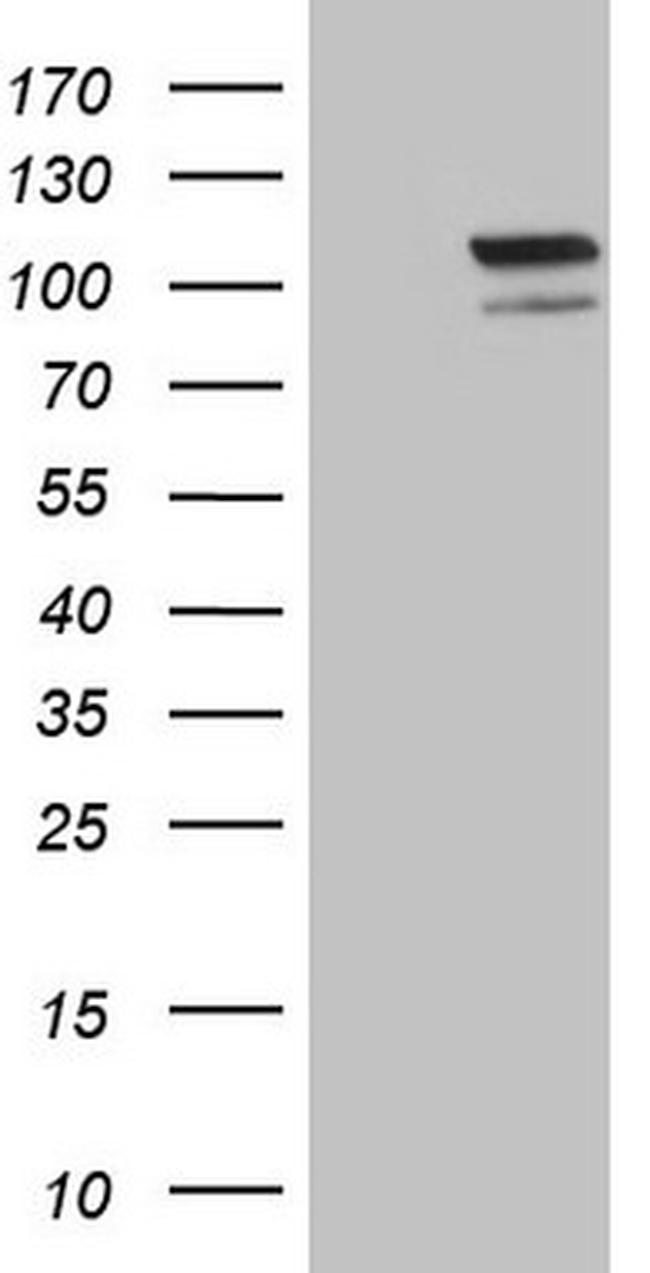
L'EPAS1 (en anglais Endothelial PAS domain-containing protein 1) ou HIF-2α (en anglais hypoxia-inducible factor-2alpha) est une protéine codée par le gène EPAS1 humain. C'est un facteur induit par l’hypoxie, un groupe de facteurs de transcription impliqué dans la réponse physiologique à la concentration en oxygène,,,. Le gène est actif en environnement hypoxique (à faible teneur en oxygène). Il est aussi important pour le développement du cœur et pour le maintien de l'équilibre catécholaminique nécessaire au bon fonctionnement du cœur. Des mutations conduisent souvent à l'apparition de tumeurs neuroendocrines.
Cependant, plusieurs allèles du gène EPAS1 contribuent à l'adaptation humaine à la haute altitude. Un de ces allèles, qui a été hérité des dénisoviens, est connu pour conférer une performance sportive accrue chez certaines personnes, et a donc été qualifié de « gène du super-athlète ».
Fonction
Le gène EPAS1 code une sous-unité d'un facteur de transcription impliqué dans l'activation de gènes régulés par l'oxygène et induite par la diminution de la concentration en oxygène (hypoxie). La protéine contient un domaine de dimérisation de la protéine hélice-boucle-hélice ainsi qu'un domaine que l’on retrouve dans les protéines de transduction du signal de réponse au niveau d'oxygène. EPAS1 est impliqué dans le développement embryonnaire du cœur et est exprimé dans cellules endothéliales qui tapissent les parois des vaisseaux sanguins dans le cordon ombilical.
EPAS1 est également essentiel pour le maintien de l'homéostasie de la catécholamine et la protection contre l'insuffisance cardiaque au début du développement embryonnaire. Les catécholamines régulées par l'EPAS1 comprennent l'épinéphrine et la norépinéphrine. Il est essentiel que la production de catécholamines reste dans des conditions homéostatiques afin que le cœur vulnerable du fœtus et le cœur de l'adulte ne s’épuisent pas et ne provoquent pas d'insuffisance cardiaque. La production de catécholamine dans l'embryon est liée au contrôle du débit cardiaque en augmentant la fréquence cardiaque fœtale.
Allèles
Un pourcentage élevé de Tibétains porte un allèle d'EPAS1 qui améliore le transport d'oxygène. L'allèle bénéfique se trouve également dans le génome des Dénisoviens, suggérant qu'il est apparu chez eux et a été transmis aux humains modernes par hybridation.
De même, le dogue du Tibet a hérité d'un allèle du gène d'adaptation à l'altitude, issu d'un métissage avec le loup tibétain.
Signification clinique
Le gène EPAS1 code le facteur de transcription HIF-2α, qui stimule la production de globules rouges et augmente ainsi la concentration d'hémoglobine dans le sang. Certaines mutations qui augmentent son expression sont associées à un accroissement des cas d'hypertension, d'accidents vasculaires cérébraux, mais aussi de cancers.
Des mutations du gène EPAS1 sont liées à l'apparition précoce de tumeurs neuroendocrines telles que les paragangliomes, les somatostatinomes ou les phéochromocytomes. Les mutations sont généralement sans conséquences somatiques et se localisent dans le site d'hydroxylation primaire de l'HIF-2α, qui perturbent le mécanisme d'hydroxylation / dégradation des protéines et conduisent à la stabilisation des protéines et des signaux pseudohypoxiques. De plus, ces tumeurs neuroendocrines libèrent de l'érythropoïétine (EPO) dans la circulation sanguine et entrainent une polycythémie, entrant dans le cadre d'un syndrome de Pacak-Zhuang.
Des mutations de ce gène sont également associées à une érythrocytose familiale de type 4, à l'hypertension pulmonaire et au mal chronique des montagnes.
À haute altitude, l'EPAS1 peut provoquer une production excessive de globules rouges, générant un mal chronique des montagnes engageant les performances et la survie. Elle y est donc soumise à une pression sélective. C'est ainsi que les allèles associés à des concentrations d'hémoglobine plus faibles s'observent à des fréquences nettement plus élevées chez les Tibétains vivant à plus de 4 000 m d’altitude par comparaison avec les populations voisines. Ceci indique que certaines variantes de ce gène protègent les personnes vivant à haute altitude comme au Tibet,,. L'effet est le plus marqué chez les Tibétains vivant dans l'Himalaya à une altitude d'environ 4 000 mètres, dont l'environnement est intolérable pour les autres populations humaines du fait d'une baisse de 40 % du niveau d'oxygène atmosphérique.
Il existe plus de 30 facteurs génétiques — dont l'EPAS1 — qui rendent l'organisme des Tibétains mieux adaptés aux hautes altitudes.
Interactions
L'EPAS1 interagit avec le translocateur nucléaire du récepteur des hydrocarbures aryliques (en) et l'ARNTL (en).
Le VHL inhibe l'EPAS1. En cas de mutation du premier, inactivant celui ci, l'activité de l'EPAS1 augmente quel que soit le niveau d'oxygène et favorisant certains cancers, dont celui du cancer du rein.
Cible thérapeutique
Le belzutifan est un inhibiteur de l'EPAS1 et a été utilisé dans le traitement du syndrome de Pacak-Zhuang.
Notes et références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « EPAS1 » (voir la liste des auteurs).
Voir aussi
Bibliographie
- Brahimi-Horn MC, Pouysségur J, « The hypoxia-inducible factor and tumor progression along the angiogenic pathway », International Review of Cytology, vol. 242, , p. 157–213 (ISBN 9780123646460, PMID 15598469, DOI 10.1016/S0074-7696(04)42004-X)
- Haase VH, « Hypoxia-inducible factors in the kidney », American Journal of Physiology. Renal Physiology, vol. 291, no 2, , F271-81 (PMID 16554418, PMCID 4232221, DOI 10.1152/ajprenal.00071.2006)
- Andersson B, Wentland MA, Ricafrente JY, Liu W, Gibbs RA, « A "double adaptor" method for improved shotgun library construction », Analytical Biochemistry, vol. 236, no 1, , p. 107–13 (PMID 8619474, DOI 10.1006/abio.1996.0138)
- Yu W, Andersson B, Worley KC, Muzny DM, Ding Y, Liu W, Ricafrente JY, Wentland MA, Lennon G, Gibbs RA, « Large-scale concatenation cDNA sequencing », Genome Research, vol. 7, no 4, , p. 353–8 (PMID 9110174, PMCID 139146, DOI 10.1101/gr.7.4.353)
- Ema M, Taya S, Yokotani N, Sogawa K, Matsuda Y, Fujii-Kuriyama Y, « A novel bHLH-PAS factor with close sequence similarity to hypoxia-inducible factor 1alpha regulates the VEGF expression and is potentially involved in lung and vascular development », Proceedings of the National Academy of Sciences of the United States of America, vol. 94, no 9, , p. 4273–8 (PMID 9113979, PMCID 20712, DOI 10.1073/pnas.94.9.4273)
- Hogenesch JB, Gu YZ, Jain S, Bradfield CA, « The basic-helix-loop-helix-PAS orphan MOP3 forms transcriptionally active complexes with circadian and hypoxia factors », Proceedings of the National Academy of Sciences of the United States of America, vol. 95, no 10, , p. 5474–9 (PMID 9576906, PMCID 20401, DOI 10.1073/pnas.95.10.5474)
- Takahata S, Sogawa K, Kobayashi A, Ema M, Mimura J, Ozaki N, Fujii-Kuriyama Y, « Transcriptionally active heterodimer formation of an Arnt-like PAS protein, Arnt3, with HIF-1a, HLF, and clock », Biochemical and Biophysical Research Communications, vol. 248, no 3, , p. 789–94 (PMID 9704006, DOI 10.1006/bbrc.1998.9012)
- Ema M, Hirota K, Mimura J, Abe H, Yodoi J, Sogawa K, Poellinger L, Fujii-Kuriyama Y, « Molecular mechanisms of transcription activation by HLF and HIF1alpha in response to hypoxia: their stabilization and redox signal-induced interaction with CBP/p300 », The EMBO Journal, vol. 18, no 7, , p. 1905–14 (PMID 10202154, PMCID 1171276, DOI 10.1093/emboj/18.7.1905)
- Cockman ME, Masson N, Mole DR, Jaakkola P, Chang GW, Clifford SC, Maher ER, Pugh CW, Ratcliffe PJ, Maxwell PH, « Hypoxia inducible factor-alpha binding and ubiquitylation by the von Hippel-Lindau tumor suppressor protein », The Journal of Biological Chemistry, vol. 275, no 33, , p. 25733–41 (PMID 10823831, DOI 10.1074/jbc.M002740200)
- Maemura K, de la Monte SM, Chin MT, Layne MD, Hsieh CM, Yet SF, Perrella MA, Lee ME, « CLIF, a novel cycle-like factor, regulates the circadian oscillation of plasminogen activator inhibitor-1 gene expression », The Journal of Biological Chemistry, vol. 275, no 47, , p. 36847–51 (PMID 11018023, DOI 10.1074/jbc.C000629200)
- Luo JC, Shibuya M, « A variant of nuclear localization signal of bipartite-type is required for the nuclear translocation of hypoxia inducible factors (1alpha, 2alpha and 3alpha) », Oncogene, vol. 20, no 12, , p. 1435–44 (PMID 11313887, DOI 10.1038/sj.onc.1204228)
- Woods SL, Whitelaw ML, « Differential activities of murine single minded 1 (SIM1) and SIM2 on a hypoxic response element. Cross-talk between basic helix-loop-helix/per-Arnt-Sim homology transcription factors », The Journal of Biological Chemistry, vol. 277, no 12, , p. 10236–43 (PMID 11782478, DOI 10.1074/jbc.M110752200)
- Lando D, Peet DJ, Whelan DA, Gorman JJ, Whitelaw ML, « Asparagine hydroxylation of the HIF transactivation domain a hypoxic switch », Science, vol. 295, no 5556, , p. 858–61 (PMID 11823643, DOI 10.1126/science.1068592)
- Mole DR, Pugh CW, Ratcliffe PJ, Maxwell PH, « Regulation of the HIF pathway: enzymatic hydroxylation of a conserved prolyl residue in hypoxia-inducible factor alpha subunits governs capture by the pVHL E3 ubiquitin ligase complex », Advances in Enzyme Regulation, vol. 42, , p. 333–47 (PMID 12123724, DOI 10.1016/S0065-2571(01)00037-1)
- Sivridis E, Giatromanolaki A, Gatter KC, Harris AL, Koukourakis MI, « Association of hypoxia-inducible factors 1alpha and 2alpha with activated angiogenic pathways and prognosis in patients with endometrial carcinoma », Cancer, vol. 95, no 5, , p. 1055–63 (PMID 12209691, DOI 10.1002/cncr.10774)
- Elvert G, Kappel A, Heidenreich R, Englmeier U, Lanz S, Acker T, Rauter M, Plate K, Sieweke M, Breier G, Flamme I, « Cooperative interaction of hypoxia-inducible factor-2alpha (HIF-2alpha ) and Ets-1 in the transcriptional activation of vascular endothelial growth factor receptor-2 (Flk-1) », The Journal of Biological Chemistry, vol. 278, no 9, , p. 7520–30 (PMID 12464608, DOI 10.1074/jbc.M211298200)
- Sang N, Stiehl DP, Bohensky J, Leshchinsky I, Srinivas V, Caro J, « MAPK signaling up-regulates the activity of hypoxia-inducible factors by its effects on p300 », The Journal of Biological Chemistry, vol. 278, no 16, , p. 14013–9 (PMID 12588875, PMCID 4518846, DOI 10.1074/jbc.M209702200)
- Portail de la médecine
- Portail de la biochimie